
Analýzy, komentáře
Smazávání rozdílu mezi dnem a nocí ovlivňuje cirkadiánní systém organismů
....ale proč by měly cirkadiánní hodiny zajímat ochranu přírody?Cirkadiánní hodiny
Cirkadiánní systém se vyvinul u všech organismů - od jednobuněčných řas po člověka - jako adaptace na 24h solární cyklus, tedy střídání dne a noci. V jaderných buňkách většiny organismů je založen na zpětnovazebných smyčkách, kterými proteiny tzv. hodinových genů blokují svojí vlastní transkripci. Hodinové geny v mnohobuněčných organismech řídí expresi více než 10 % genů zapojených do regulace buněčného cyklu, metabolismu, imunitních či regeneračních procesů v organismu a další cirkadiánní regulace probíhají na úrovni proteinových modifikací. Velké procento proteomu (soubor proteinů celého organismu) je tak ekonomicky regulováno v souladu se spánkovým rytmem.
Ačkoliv jsou cirkadiánní hodiny většiny živočichů schopné chodu i ve stálé tmě, nefungují přesně a potřebují každodenní kalibraci neboli synchronizaci se solárním cyklem. Z humánní medicíny a preklinických laboratorních experimentů na hlodavcích víme, že narušení této synchronizace vysoce zvyšuje riziko vzniku celé řady nemocí, od různých forem depresí, přes diabetes 2. typu, zánětlivá onemocnění, hypertenzi až po rakovinu.
Synchronizace se solárním 24h cyklem je morfologicky zajištěna propojením centrálních cirkadiánních pacemakerů (udavatelů rytmu, podobných pacemakerům v srdci, které udávají srdeční frekvenci, jen pracující s cirkadiánní periodou) v mozku se sítnicí oka a, pokud nemluvíme pouze o savcích, i s dalšími světločivnými orgány, jako je pineální žláza ptáků, plazů apod. Pacemakery potom posílají informaci o přesné denní době do celého organismu neuronálními a humorálními signály. Jedním z nejdůležitějších takových signálů je rytmická produkce pineálního hormonu melatoninu. Jeho syntéza je regulována centrálním pacemakerem tak, aby probíhala v noci. Melatonin se vylévá do krevního oběhu a má receptory ve všech orgánech těla. Takto signalizuje periferním hodinám v buňkách noční fázi 24h cyklu, a funkční proteom v jednotlivých orgánech si tím zachovává přesný fázový vztah se střídáním světla a tmy ve vnějším prostředí.
Vývoj cirkadiánního systému denních i nočních živočichů má z evolučního pohledu stejný základ. Noční tvorové nejsou vystaveni celé fotoperiodě, a proto se jako nejefektivnější způsob synchronizace ukázalo vytvoření časového „okna“ citlivosti ke světlu, které se otevírá vždy před a po periodě tmy, což v přírodě znamená soumrak a svítání. Čím větší kontrast mezi světlem a tmou předchází či následuje po tomto „oknu“, tím přesnější je synchronizace cirkadiánních hodin se solárním cyklem a také tím přesnější je synchronizace jednotlivých orgánových soustav mezi sebou.
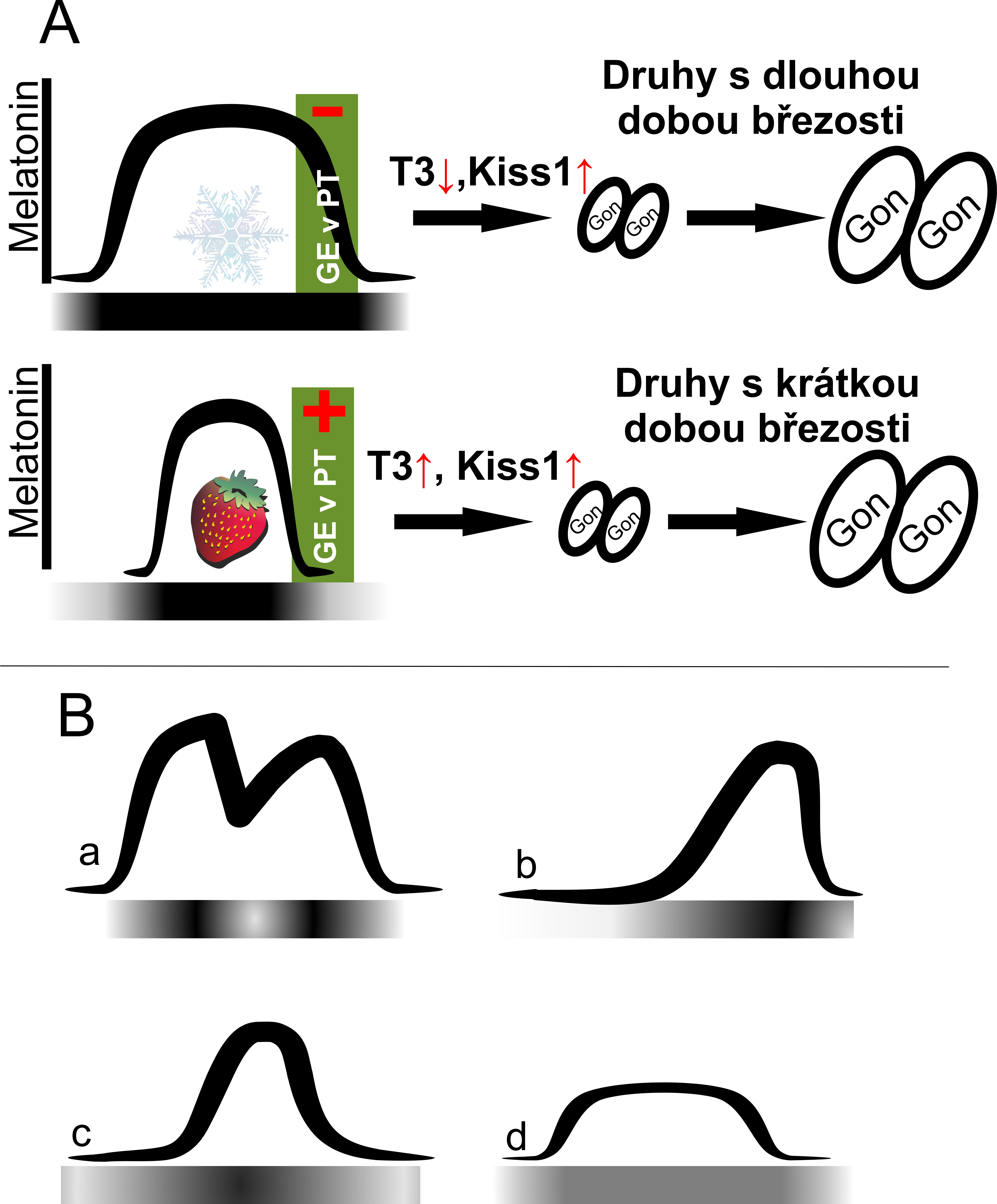
Světelná synchronizace cirkadiánního systému není podstatná jen pro udržování cirkadiánních rytmů, je to také základní mechanismus, který umožňuje adaptaci organismů k zeměpisné šířce a sezónním změnám v přírodě. Měnící se poměr během dne a noci v průběhu roku se kopíruje do nastavení fází cirkadiánních pacemakerů a poté do vlnové formy cirkadiánních rytmů. Letní komprese a zimní dekomprese vysoké noční hladiny melatoninu je nezastupitelný signál pro savce i ptáky pro spuštění sezónních procesů v organismech (viz obr. 1A).
Světlo v noci
Antropogenní světlo v noci má dva hlavní dopady na cirkadiánní a sezónní fyziologii. Jednak komplikuje synchronizaci k solárnímu 24h cyklu tím, že snižuje kontrast mezi dnem a nocí a snižuje tak rozlišitelnost mezi dnem a nocí pro cirkadiánní systém. Toto je v dnešní době jasně popsaný problém pro živočichy v městském prostředí, a ve volné přírodě to znamená ohrožení zejména pro noční živočichy, kteří jsou vnímaví k velmi nízkým intenzitám světla signalizujícím vlastně konec soumraku a začátek svítání. Díky rychlému trendu zvyšování intenzity nočního světla v posledních letech a rychlosti jeho šíření do volné přírody z měst díky závojovému jasu (v angl. „skyglow) lze očekávat, že se problém bude týkat brzy všech živočichů ve volné krajině včetně člověka.
Světlo v noci také okamžitě, v řádu několika minut, snižuje noční hladinu melatoninu (obr. 1B). Chronicky snížená hladina melatoninu má za následek komplikace v sezónních cyklech reprodukce, migrace, hibernace nebo jiných adaptačních změnách ve fyziologii. Nízká hladina melatoninu ale také narušuje imunitní a metabolické procesy a rytmus spánku a bdění, a to jak u nočních, tak u denních živočichů.
Ačkoliv základní procesy cirkadiánních regulací jsou shodné u denních a nočních živočichů, z laboratorních experimentů je zjevné, že citlivost melatoninu nočních živočichů na světlo v noci je výrazně vyšší. Jedny z prvních studií ze 70. a začátku 80. let ukázaly, že 8 minutová expozice bílému světlu o intenzitě nižší než 1 lux potlačuje u syrských křečků do 10 minut hladinu melatoninu o více než 25 % a melatonin zůstává nízký, i když jsou zvířata znovu vrácena do tmy1. Podobný účinek 1 minutové expozice světlu popsala také prof. Illnerová u potkanů2. Jasné světlo, kolem 1000 luxů, může dokonce působit na křečka jen 1 vteřinu a hladina jeho melatoninu spolehlivě klesne3. Noční světlo u hlodavců vede dokonce ke vzniku jakési „paměti“. Ukázalo se totiž, že krátké 1 minutové světlo nesníží melatonin jen v době svého působení a krátce nato, ale melatonin je ve stejnou dobu snížen i v několika následujících cyklech4. Vysokou citlivost na noční světlo mají také ryby a žáby, které žijí ve velmi tmavém nočním prostředí, a jimž stačí intenzita světla nižší než intenzita měsíce v úplňku k významnému potlačení noční hladiny melatoninu5.
Citlivost denních živočichů k nočnímu světlu je řádově vyšší. Nicméně 10-30 luxů snižujících melatonin u japonských makaků (Macaca fuscata fuscata) je na úrovni intenzity běžného osvětlení městského prostředí6. Velmi častým modelem denních živočichů studovaným v souvislosti se světlem v noci jsou ptáci. Nedávná studie ukázala, že již 1,5 luxu stačí zebřičkám pestrým (Taeniopygia guttata) k potlačení melatoninu a dokonce 0,3 luxu, tedy světlo měsíce v úplňku, stačí pro změnu v hladině melatoninu u kosa černého (Turdus merula)7,8. Jsou-li tito ptáci chováni v prostředí, kde je 0,3 luxu v noci trvale v průběhu roku, nenastane u nich v dalším roce reprodukční fáze a nezvýší se zjara hladina jejich testosteronu.
V posledních letech roste počet prací, které dokazují, že načasování sezónních reprodukčních procesů se liší mezi obratlovci obývajícími oblasti se světelným znečištěním a stejnými druhy obývajícími oblasti s malým nebo žádným antropogenním světlem. Vzhledem k nezastupitelné roli melatoninu v regulaci sezónní reprodukce, může jeho chronické potlačení (obr. 1B a,d) nebo maskování změn v přirozené délce dne (obr. 1B b,c), posunout nebo dokonce inhibovat reprodukční zrání a rozmnožování velké skupiny zvířat. Studie zabývající se touto otázkou zjistily, že ptáci žijící v městském prostředí s vysokou intenzitou nočního světla zahajují sezónní reprodukční procesy dříve než ptáci v tmavých oblastech. Tato mláďata však mají často menší šanci na přežití díky nejisté dostupnosti potravy a suboptimálním teplotám9. Také laboratorní experimenty se savci prokázaly, že 5 luxů světla během noční fáze je dostatečné k narušení fyziologických reakcí na zkracující se den, jako je houstnutí či změna barvy srsti, aktivita pohlavních žláz či nárůst tělesné hmotnosti. Tato zvířata měla také horší imunitní reakci k bakteriální infekci, což svědčí o špatné cirkadiánní synchronizaci obdobné, jako známe u lidí dlouhodobě pracujících ve směnných provozech10.
Problémy s nočním světlem jsou stále intenzivněji vnímány všude na světě. Z jedné práce publikované na téma „light at night AND ecosystems“ v 60. letech se počet prací na toto téma vyšplhal na více než 400 v posledním desetiletí (zdroj: PubMed). Dlouhodobé sledování klokanů dama (Macropus eugenii) v Austrálii např. ukázalo, že zvířata žijících v oblastech s vysokým světelným znečištěním rodí později a vrhy mláďat jsou méně synchronizovány s vhodným ročním obdobím než vrhy klokanů žijících v oblastech bez světelného znečištění. Autoři naznačují, že antropogenní světlo maskuje malé rozšíření přirozené noci po letním slunovratu, které klokani využívají k reaktivaci blastocyst11. Jiná práce referuje o tom, že samci makiho trpasličího (Microcebus murinus) vystavení nočnímu světlu simulujícímu intenzitu a spektrum pouličních světel v Brunoy ve Francii, nastupují do říje o 2 týdny dříve než u samci vystavení měsíčnímu světlu, díky změnám v hladině melatoninu12. Tyto a mnoho dalších studií ukazují, že antropogenní světlo v noci může ovlivnit načasování sezónní reprodukce a vyvolat tak problémy s udržením populací mnoha živočišných, nejen nočních, druhů.
Modré světlo
V posledních letech se často mluví o faktoru tzv. modrého světla, které je v minimální míře součástí spektra zastaralých sodíkových výbojek, má ale velmi významný podíl ve spektru nových LED svítidel, která sodíkové výbojky celosvětově nahrazují. Obavy z rostoucího podílu modrého spektra ve veřejném osvětlení a reklamních či jiných světelných zdrojích vycházejí ze základního principu vnímání světla zrakovou dráhou vedoucí k cirkadiánním pacemakerům, která je morfologicky odlišná od dráhy pro obrazové vidění. Tato dráha je „vyladěná“ tak, že s velkou citlivostí vnímá světelné spektrum v blankytné modré, okolo vlnových délek 480 nm. Obsahuje speciální fotopigment melanopsin, který byl objeven zhruba před pětadvaceti lety, tj. přibližně v době, kdy se, spolu se zaváděním světelných zdrojů LED do veřejného prostoru, začaly objevovat první problémy s dopadem na ekosystém. Spektrální složení prvních LED svítidel se totiž překrývá se spektrální citlivostí melanopsinu a tím i cirkadiánního systému, a tak i systému řídícího sezónní fyziologii. Postupem času se ukázalo, že vyladění k „modré“ není vlastností jen obratlovců, ale že fotopigment s těmito vlastnostmi existuje u většiny živočichů včetně hmyzu.
Z hlediska kompromisu mezi nároky zrakového systému člověka, jediného uživatele nočního osvětlování na planetě, a zachování rozmanitosti přírody a zdraví jednotlivých živočichů, se jeví jako ideální moderní trend zavádění biodynamického osvětlování. To se řídí solárním cyklem a cyklem aktivity člověka a tlumí intenzitu a mění spektrum v době, kdy není pro člověka bezprostředně užitečné. Existuje však řada organismů, zejména rostlin a hmyzu, ale pravděpodobně i některých druhů ptáků a savců, které jsou citlivé i k červenému či zelenému světlu. Také zcela neplatí, že světlo jiných spekter než je modrá, uniká zcela pozornosti cirkadiánním hodinám. Musí být jenom trochu silnější k vyvolání stejného účinku. Je proto mít na paměti, že i moderní technologie svícení ve veřejném prostoru jsou stále pouze kompromisem a jedinou možnou prevencí proti vymazání nočního prostředí je osvěta a cílená smysluplná regulace zavádění nových světelných zdrojů per se.
Literatura:
1Brainard GC, Richardson BA, Hurlbut EC, Steinlechner S, Matthews SA, Reiter RJ. (1984). The influence of various irradiances of artificial light, twilight, and moonlight on the suppression of pineal melatonin content in the Syrian hamster. Journal of Pineal Research, 1: 105-119. doi: 10.1111/j.1600-079x.1984.tb00202.x.
2 Illnerová H, Vanĕcek J. (1979). Response of rat pineal serotonin N-acetyltransferase to one min light pulse at different night times. Brain Research, 167: 431-434. doi: 10.1016/0006-8993(79)90841-2.
3 Reiter RJ, Joshi BN, Heinzeller T, Nürnberger F. (1986). A Single 1- or 5-Second Light Pulse at Night Inhibits Hamster Pineal Melatonin. Endocrinology, 118: 1906-1909. doi: 10.1210/endo-118-5-1906.
4 Lerchl A. (1995). Sustained response of pineal melatonin synthesis to a single one-minute light pulse during night in Djungarian hamsters (Phodopus sungorus). Neuroscience Letters, 198: 65-67. doi: 10.1016/0304-3940(95)11952-s.
5 Kupprat F, Hölker F, Kloas W. (2020). Can skyglow reduce nocturnal melatonin concentrations in Eurasian perch? Environ Pollut, 262:114324. doi: 10.1016/j.envpol.2020.114324.
6 Nozaki M, Tsushima M, Mori Y. (1990). Diurnal changes in serum melatonin concentrations under indoor and outdoor environments and light suppression of nighttime melatonin secretion in the female Japanese monkey. Journal of Pineal Research, 9: 221-230. doi: 10.1111/j.1600-079x.1990.tb00710.x.
7 Moaraf S, Vistoropsky Y, Pozner T, Heiblum R, Okuliarová M, Zeman M, Barnea A. (2020). Artificial light at night affects brain plasticity and melatonin in birds. Neurosci Lett. 716:134639. doi: 10.1016/j.neulet.2019.134639.
8 Dominoni DM, Quetting M, Partecke J. (2013). Long-Term Effects of Chronic Light Pollution on Seasonal Functions of European Blackbirds (Turdus merula). PLOS ONE 8(12): e85069. https://doi.org/10.1371/journal.pone.0085069
9 Kempenaers B, Borgström P, Loës P, Schlicht E, Valcu M. (2010). Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Current Biology, 20: 1735-1739. doi: 10.1016/j.cub.2010.08.028.
10 Aubrecht TG, Weil ZM, Nelson RJ. (2014). Dim light at night interferes with the development of the short-day phenotype and impairs cell-mediated immunity in Siberian hamsters (Phodopus sungorus). J Exp Zool A Ecol Genet Physiol. 321(8):450-456. doi: 10.1002/jez.1877.
11 Robert KA, Lesku JA, Partecke J, Chambers B. (2015). Artificial light at night desynchronizes strictly seasonal reproduction in a wild mammal. Proceedings of the Royal Society B, Biological Sciences, 282: 1-7. doi: 10.1098/rspb.2015.1745.
12 Le Tallec T, Perret M, Théry M.( 2016). Melatonin concentrations and timing of seasonal reproduction in male mouse lemurs (Microcebus murinus) exposed to light pollution. Journal of Mammalogy, 97: 753–760. https://doi.org/10.1093/jmammal/gyw003
Další čtení:
Falcón J, Torriglia A, Attia D, Viénot F, Gronfier C, Behar-Cohen F, Martinsons C, Hicks D. (2020). Exposure to Artificial Light at Night and the Consequences for Flora, Fauna, and Ecosystems. Front Neurosci, 14:602796. doi: 10.3389/fnins.2020.602796.
Bendová Z, Moravcová S. (2018). Erasing day/night differences in light intensity and spectrum affect biodiversity and the health of mammals by confusing the circadian clock. Lynx, new series. Prague: National Museum, 49(1), 139–161. DOI: https://doi.org/10.2478/lynx-2018-0012. Le Tallec T, Perret M, Théry M.
Diskuze:
V diskuzi nejsou žádné příspěvky.Pro přidávání komentářů je nutné se přihlásit nebo zaregistrovat.